

译作分享
译者:李梅
Copper and Zinc, Biological Role and Significance of Copper/Zinc Imbalance
铜和锌,铜/锌失衡的生物学作用和意义
1. UniversityMedical Center Ljubljana, Zaloska cesta 2, 1000 Ljubljana, Slovenia
2. GeneralHospital Izola, Polje40,6310 Izola, Slovenia
Introduction/介绍
The human body has an elaborate system for managing and regulating the amount of key trace metals circulating in blood and stored in cells. Nutrient metals from our diet are incorporated into blood if blood levels are depleted, transported into cells if cellular levels are inadequate, or excreted if blood and cell levels are sufficient or overloaded. When this system fails to function properly, abnormal levels and ratios of trace metals can develop. One of the most common trace-metal imbalances is elevated copper and depressed zinc. The ratio of copper to zinc is clinically more important than the concentration of either of these trace metals [1].
人体有一套精细的系统管理和调节血液循环及储存在细胞中关键一定数量的微量元素。如果血液水平耗尽,我们饮食中的营养素会被吸收到血液中,如果细胞水平不足,则会被输送到细胞中,如果血液和细胞水平充足或超负荷,则会被排出体外。当该系统无法正常运行时,微量金属的水平和比例可能会出现异常。最常见的微量金属元素失衡之一是铜含量升高和锌含量降低。在临床上铜与锌的比例比这些微量金属各自的浓度更重要 [1]。
There are 2-4 grams of Zn distributed throughout the human body [2]. Most zinc is in the brain, muscle, bones,kidney and liver, with the highest concentrations in the prostate and parts of the eye [3]. It is the second most abundant transition metal in organisms after iron and it is the only metal which appears in all enzyme classes [2,4]. Copper is also a vital dietary nutrient, although only small amounts of the metal are needed for well-being [5]. Although copper is the third most abundant trace metal in the body [behind iron and zinc], the total amount of copper in the body is only 75-100 milligrams [6]. Copper is present in every tissue of the body, but is stored primarily in the liver, with fewer amounts found in the brain, heart, kidney, and muscles [7].
人体中分布有2-4克的锌 [2]。大多数锌存在于大脑、肌肉、骨骼、肾脏和肝脏中,在前列腺和部分眼睛中的浓度最高 [3]。它是生物体中仅次于铁的第二丰富的过渡金属,也是唯一出现在所有酶类中的金属 [2,4]。尽管健康只需要少量的金属,铜也是一种重要的膳食营养素[5]。虽然铜是体内含量第三丰富的微量金属 [仅次于铁和锌],但体内铜的总量仅为75-100毫克 [6]。铜存在于身体的每个组织,但主要储存在肝脏中,在大脑、心脏、肾脏和肌肉中的含量较少 [7]。
The Path and the Role of Zinc [Zn] in Organisms/锌【Zn】在生物体中的途径和作用
Biological role of Zn/锌的生物学作用
Zinc is involved in numerous aspects of cellular metabolism [8]. It was estimated that about 10% of human proteins potentially bind zinc, in addition to hundreds which transport and traffic zinc. It is required for the catalytic activity of more than 200 enzymes [9,10] and it plays a role in immune function [10,11], wound healing [10], protein synthesis, DNA synthesis and cell division [12]. Zinc is required for proper sense of taste and smell [13,14] and supports normal growth and development during pregnancy, childhood, and adolescence [15-18]. It is believed to possess antioxidant properties, which may protect against accelerated aging and helps speed up the healing process after an injury; however, studies differ as to its effectiveness [16,19]. Zinc ions are effective antimicrobial agents even at low concentrations. Gastroenteritis is strongly attenuated by ingestion of zinc and this effect could be due to direct antimicrobial action of the zinc ions in the gastrointestinal tract, or to the absorption of the zinc and re-release from inmune cells [all granulocytes secrete zinc], or both [20,21,22]. Cells in the salivary gland, prostate, immune system and intestine use Zn signaling as one way to communicate with other cells [23]. In the brain, zinc is stored in specific synaptic vesicles by glutamatergic neurons and can modulate brain excitability [24]. It plays a key role in synaptic plasticity and so in learning [24,26]. It also can be a neurotoxin, suggesting zinc homeostasis plays a critical role in normal functioning of the brain and central nervous system [24].
锌参与许多方面的细胞代谢 [8]。据估计,除了数百种运输和转运锌的蛋白质外,大约 10% 的人类蛋白质可能与锌结合。它是200多种酶的催化活性所必需的 [9,10],它在免疫功能 [10,11]、伤口愈合 [10]、蛋白质合成、DNA 合成和细胞分裂 [12] 中发挥作用。锌是正常味觉和嗅觉所必需的 [13,14],并支持怀孕、儿童和青春期的正常生长和发育 [15-18]。它被认为具有抗氧化特性,可以防止加速老化,并有助于加速受伤后的愈合过程,然而,关于其有效性的研究存在差异 [16,19]。锌即使在低浓度下,离子也是有效的抗菌剂。摄入锌可大大减轻胃肠炎,这效果可能是由于锌离子在胃肠道中的直接抗菌作用,或锌的吸收和免疫细胞重新释放 [所有粒细胞都分泌锌],或两者兼而有之 [20,21,22]。唾液腺、前列腺、免疫系统和肠道中的细胞使用锌信号作为与其他细胞交流的一种方式 [23]。在大脑中,锌被谷氨酸能神经元储存在特定的突触小泡中,可以调节大脑的兴奋性 [24]。它在突触可塑性等学习中起着关键作用 [24-26]。它也可能是一种神经毒素,表明锌稳态在大脑和中枢神经系统的正常功能中起着关键作用 [24]。
Zn metabolism/锌代谢
In blood plasma, Zn is bound to and transported by albumin (60%, low-affinity) and transferrin (10%) [27]. Since transferrin also transports iron, excessive iron can reduce zinc absorption, and vice-versa [28]. The concentration of zinc in blood plasma stays relatively constant regardless of zinc intake [29]. Zinc may be held in metallothionein reserves and also transferred in metal transporters of ZIP and ZnT family transporter proteins [30]. Metallothioneins in intestinal cells are capable of adjusting absorption of zinc by 15– 40% [31]. Excess zinc particularly impairs copper absorption because metallothioneins absorb both metals [32].
在血浆中,锌与白蛋白(60%,低亲和力)和转铁蛋白(10%)结合并由其转运[27]。由于转铁蛋白也转运铁,过量的铁会降低锌的吸收,反之亦然 [28]。无论锌摄入量如何,血浆中锌的浓度都保持相对恒定 [29]。锌可以保留在金属硫蛋白储备中,也可以在ZIP和ZnT家族转运蛋白的金属转运蛋白中转移 [30]。肠细胞中的金属硫蛋白能够将锌的吸收调整15-40% [31]。过量的锌会损害铜的吸收,因为金属硫蛋白会吸收两种金属 [32]。
Zn enzymes/锌酶
Two examples of zinc-containing enzymes are carbonic anhydrase and carboxypeptidase, which are vital to the processes of carbon dioxide (CO2 )regulation and digestion of proteins, respectively [33,34]. In vertebrate blood, carbonic anhydrase (Figure 1) converts CO2 into bicarbonate and the same enzyme transforms the bicarbonate back into CO2 for exhalation through the lungs [35]. Without this enzyme, this conversion would occur about one million times slower at the normal blood pH of 7 or would require a pH of 10 or more [36]. Carboxypeptidase cleaves peptide linkages during digestion of proteins. A coordinate covalent bond is formed between the terminal peptide and a C=O group attached to zinc, which gives the carbon a positive charge. This helps to create a hydrophobic pocket on the enzyme near the Zn, which attracts the non-polar part of the protein being digested [34]. Zn serves a purely structural role in zinc fingers (Figure 2) [37]. Zinc fingers form parts of some transcription factors, which are proteins that recognize DNA base sequences during the replication and transcription of DNA. Each of the nine or ten Zn2+ ions in a zinc finger helps maintain the finger’s structure by coordinately binding to four amino acids in the transcription factor [38]. The transcription factor wraps around the DNA helix and uses its fingers to accurately bind to the DNA sequence [39,40]. Zn ions are coordinated to the amino acid side chains of aspartic acid, glutamic acid, cysteine and histidine [41]. The metal also has a flexible coordination geometry, which allows proteins using it to rapidly shift conformations to perform biological reactions [42].
含锌酶的两个例子是碳酸酐酶和羧肽酶,它们分别对二氧化碳 (CO2) 的调节和蛋白质的消化过程至关重要 [33,34]。在脊椎动物血液中,碳酸酐酶(图 1)将CO2转化为碳酸氢盐,同样的酶将碳酸氢盐转化回CO2,通过肺部呼出 [35]。如果没有这种酶,这种转化在正常血液pH值为7时会慢约一百万倍,或者需要10或更高的pH值 [36]。羧肽酶在蛋白质消化过程中裂解肽键。在末端肽和附着在锌上的C=O基团之间形成配位共价键,使碳带正电荷。这有助于在锌附近的酶上形成疏水袋,从而吸引正在消化的蛋白质的非极性部分 [34]。锌在锌指中起纯粹的结构作用(图 2)[37]。锌指形成一些转录因子的一部分,这些转录因子是在DNA复制和转录过程中识别DNA碱基序列的蛋白质。锌指中的9个或10个Zn2+离子中的每一个都通过与转录因子中的四个氨基酸协同结合来帮助维持手指的结构[38]。转录因子缠绕在DNA螺旋周围,并使用其手指准确地与DNA序列结合[39,40]。锌离子与天冬氨酸、谷氨酸、半胱氨酸和组氨酸的氨基酸侧链配位[41]。这种金属还具有灵活的配位几何结构,允许蛋白质使用它快速改变构象以进行生物反应[42]。

图1:人体碳酸酐酶示意图,其中锌原子可见中心
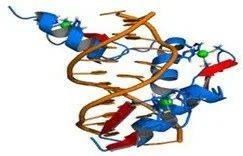
图2:由锌指中的两个组氨酸侧链协调的锌离子(绿色)
Zn transport system/锌运输系统
Zinc functions in biology are numerous, but can be separated into three main categories: catalytic, regulatory, and structural roles [42]. Greater than ten percent of the human genome codes for zinc-containing proteins [43]. Zn homeostasis is controlled by the coordinated actions of Zn transporters, which are responsible for Zn influx and efflux, and regulate the intracellular and extracellular Zn concentration and distribution. Zn transporters contribute to cellular events at the molecular, biochemical, and genetic level, with recent progress uncovering the roles of Zn transporters in physiology and pathogenesis [42,43].
Tight-control of cellular zinc homeostasis is maintained by proteins that can affect the amount of available zinc. Metal transporters of ZIP family and zinc transporters of ZnT family, as well as zinc-binding by metallothioneins; maintain control of intracellular zinc levels. Currently, 10 ZnT and 14 ZIP transporters have been identified. The ZnT proteins function in cellular zinc efflux or vesicular storage. ZnT1 was the first zinc transporter to be characterized, and is expressed in all tissues, localizing to the plasma membrane of cells [43]. Subsequent studies revealed zinc-regulated expression of ZnT1, and that zinc regulates expression of ZnT1 through activation of the metal response element – metal binding transcription factor MTF-1. Already known metal-inducible genes regulated by MTF-1 include the metallothioneins, glutamate–cysteine ligase heavy chain (which codes for an oxidative stress-related protein), and already mentioned ZnT1. MTF-1 responds to changes in intracellular zinc and other heavy-metals (cadmium and copper), where upon metal-occupancy there is translocation of MTF-1 from the cytosol to the nucleus. Subsequently this transcription factor binds to metal response elements located in the proximal promoters of metal responsive genes, resulting in an increased rate of transcription [43]. The importance of MTF-1 to zinc homeostasis and mammalian physiology is accentuated by the fact that ablation of the gene leads to severe liver degeneration and embryonic lethality in mice. Subsequent investigation of a liver-specific conditional knockout of the MTF-1 gene in mice revealed another MTF-1 regulated zinc transporter gene – ZIP10: The experiment on isolated mice hepatocytes showed that MTF- 1 is an integral part of ZIP10 related cellular zinc homeostasis in the liver both during zinc restriction and zinc excess. The results showed that MTF-1 has physiologic significance and can act as a repressor of ZIP10 under normal cellular levels of zinc. Upon reducing cellular zinc, repression is lost and MTF-1 is not translocated to the nucleus, allowing enhanced ZIP10 transcriptional activation. The results also show that ZIP10 is a new target to investigate dietary influences on zinc metabolism by the liver and within specific regions of the brain. Zinc is believed to have second messenger functions in brain and the ZIP transporters are believed to be a key to cellular Zn homeostasis [43].
锌在生物学中的功能很多,但可以分为三大类:催化作用、调节作用和结构作用 [42]。超过10%的人类基因组编码含锌蛋白质 [43]。锌的稳态受锌转运蛋白的协同作用控制,锌转运蛋白负责锌的流入和流出,并调节细胞内和细胞外锌的浓度和分布。锌转运蛋白在分子、生化和遗传水平上都发挥了作用,最近的进展揭示了锌转运蛋白在生理学和发病机制中的作用[42,43]。
对细胞锌稳态起严格控制作用的是那些可以影响可用锌量的蛋白质。ZIP家族的金属转运蛋白和ZnT家族的锌转运蛋白,和金属硫蛋白与锌结合一样,维持对细胞内锌水平的控制。目前,已鉴定出10种ZnT和14种ZIP转运蛋白。ZnT蛋白在细胞锌流出或囊泡储存中起作用。ZnT1是第一个被表达的锌转运蛋白,并在所有组织中表达,定位于细胞的质膜 [43]。随后的研究揭示了ZnT1的锌调节表达,并且锌通过激活金属反应元件——金属结合转录因子MTF-1来调节ZnT1的表达。已知的受MTF-1调节的金属诱导基因包括金属硫蛋白、谷氨酸-半胱氨酸连接酶重链(编码氧化应激相关蛋白)和已经提到的ZnT1。MTF-1回应细胞内锌和其他重金属(镉和铜)的变化,当金属占据时,MTF-1从细胞质转移到细胞核。随后,该转录因子与位于金属反应基因近端启动子中的金属反应元件结合,导致转录率增加[43]。MTF-1对锌稳态和哺乳动物生理学的重要性因基因消融导致小鼠严重肝退化和胚胎致死这一事实而更加突出。随后对小鼠MTF-1基因的肝脏特异性条件性敲除的研究揭示了另一个MTF-1调节的锌转运蛋白基因-ZIP10:对离体小鼠肝细胞的实验表明,在肝脏中,在锌限制和锌过量期间,MTF-1是ZIP10相关细胞锌稳态的组成部分。结果表明,MTF-1在正常细胞锌水平下可作为ZIP10的阻遏物,具有生理意义。减少细胞锌后,抑制作用消失,MTF-1不会转移到细胞核,允许增强的ZIP10转录激活。结果还表明,ZIP10是研究饮食对肝脏和大脑特定区域内锌代谢影响的新目标。锌被认为在大脑中具有第二信使功能,并且ZIP转运蛋白被认为是细胞锌稳态的关键因素[43]。
接Part.2
- 还没有人评论,欢迎说说您的想法!